📋 Key Information Summary
- B cells and T cells are the adaptive immune system's primary effectors; their cooperation generates high-affinity antibodies and durable immunological memory
- CD4⁺ T helper cells provide critical cognate help to B cells via CD40L–CD40 interaction, ICOS–ICOSL costimulation, and cytokine signals
- T follicular helper (Tfh) cells are the specialised CD4 subset essential for germinal centre formation and affinity maturation
- Germinal centres are organised lymphoid structures where B cells undergo somatic hypermutation and class-switch recombination to produce high-affinity, class-switched antibodies
- The germinal centre reaction separates into a dark zone (proliferation/mutation) and light zone (selection via follicular dendritic cell antigen display)
- B cell class switching is directed by cytokines: IL-4 → IgG1/IgE, IFN-γ → IgG2/IgG3, TGF-β → IgA, BAFF/APRIL → IgA at mucosal sites
- Long-lived plasma cells migrate to bone marrow survival niches and secrete antibody for years to decades after immunisation or infection
- Memory B cells and memory T cells are formed both within and outside germinal centres, providing rapid secondary responses
- Defects in B–T cell collaboration underlie primary immunodeficiencies (e.g., Hyper-IgM syndrome, CVID) and are exploited in autoimmune disease
- Australian vaccination programmes (NIP schedule) rely on intact germinal centre reactions to generate protective antibody titres
- ATSI populations experience higher rates of invasive pneumococcal disease and influenza-related hospitalisation, making understanding of humoral immunity clinically important
Introduction & Australian Context
Cooperation between B lymphocytes and T lymphocytes within germinal centres drives the generation of high-affinity antibody responses and long-lived immunological memory — the foundation of protective immunity conferred by vaccination and natural infection.
Understanding B–T cell collaboration is essential for Australian clinicians managing immunodeficiency, autoimmune disease, transplant immunology, and vaccine programmes. Australia's National Immunisation Program (NIP) schedules vaccines across the lifespan, relying on robust germinal centre responses for protection against pneumococcal disease, influenza, measles, pertussis, and SARS-CoV-2.
Primary immunodeficiency affecting B–T cell cooperation — including X-linked agammaglobulinaemia, common variable immunodeficiency (CVID), and combined immunodeficiencies — affects approximately 1 in 1,200 Australians, though many remain undiagnosed. The Australasian Society of Clinical Immunology and Allergy (ASCIA) maintains diagnostic and management guidelines for these conditions.
This guideline provides a clinical overview of B–T cell biology with emphasis on the germinal centre reaction, CD4 helper functions, and memory formation, framed for primary care physicians, immunologists, haematologists, and infectious diseases specialists practising in Australia.
B–T Cell Collaboration
The adaptive immune response requires intimate cooperation between antigen-specific B cells and CD4⁺ T helper cells. This collaboration occurs primarily in secondary lymphoid organs (lymph nodes, spleen, mucosal-associated lymphoid tissue) and is essential for the production of class-switched, high-affinity antibodies.
Antigen Presentation and T Cell Priming
The collaboration begins when dendritic cells capture antigen in peripheral tissues and migrate to draining lymph nodes. There, they present processed peptide–MHC class II complexes to naïve CD4⁺ T cells. Successful T cell activation requires three signals:
- Signal 1: TCR engagement with peptide–MHC II complex on the dendritic cell
- Signal 2: Costimulation via CD28 (T cell) binding B7-1/B7-2 (CD80/CD86) on the dendritic cell
- Signal 3: Cytokine milieu that directs T helper cell differentiation
The Immunological Synapse
Activated CD4⁺ T cells migrate to the T–B cell border of lymphoid follicles. B cells that have captured and processed antigen via their B cell receptor (BCR) present peptide–MHC II complexes to cognate T helper cells. This interaction forms an immunological synapse characterised by:
- TCR–MHC II recognition: Antigen-specific, MHC-restricted contact
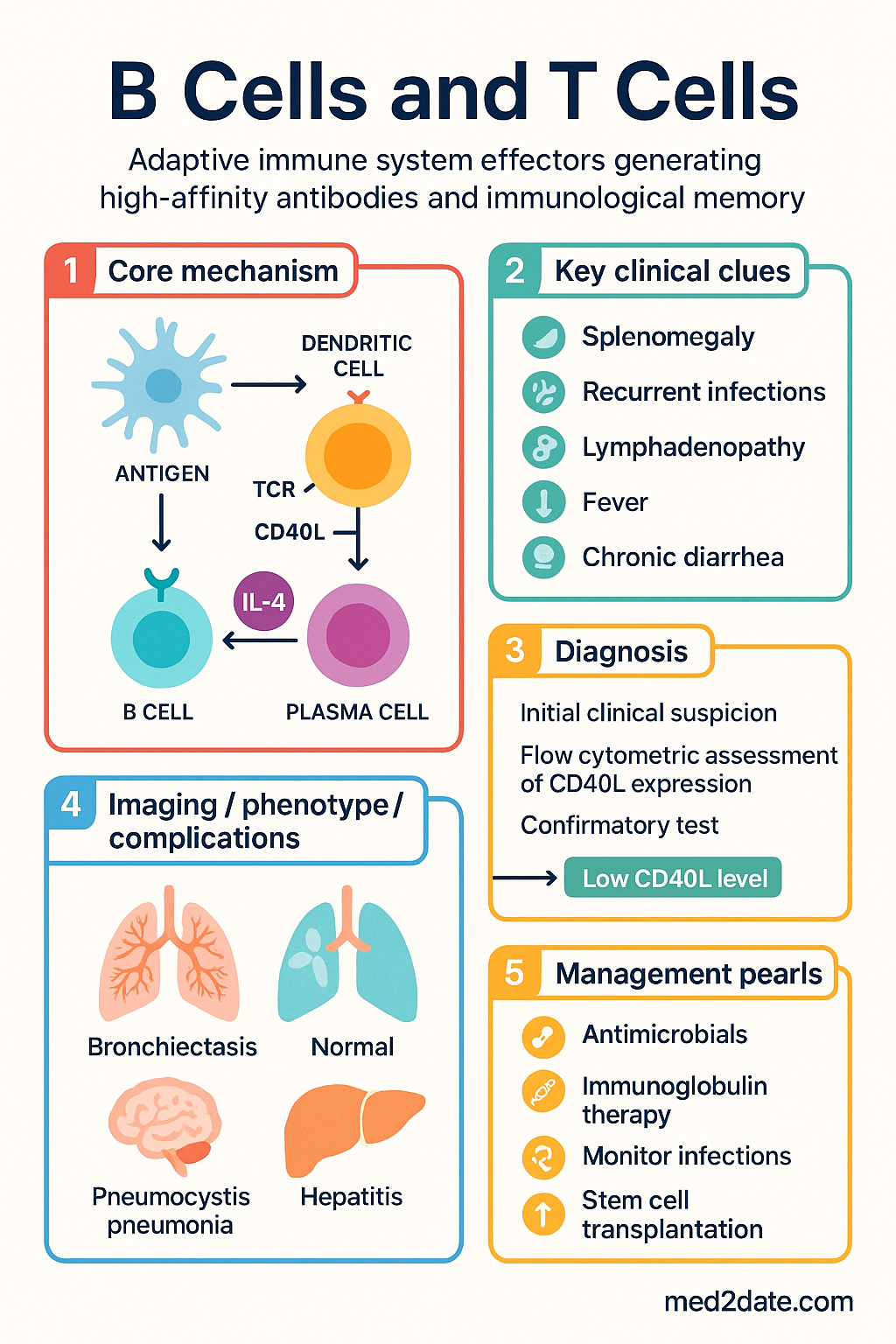
- CD40L (T cell) – CD40 (B cell): Critical costimulatory signal that rescues B cells from apoptosis and promotes proliferation, germinal centre entry, and class switching
- ICOS (T cell) – ICOSL (B cell): Essential for Tfh cell function and germinal centre maintenance
- SAP/SLAM interactions: Regulate stable T–B conjugate formation; deficiency causes X-linked lymphoproliferative disease
Thymus-Dependent vs Thymus-Independent Responses
Antigens are classified based on their T cell dependence:
| Feature | Thymus-Dependent (TD) | Thymus-Independent (TI) |
|---|---|---|
| Antigen type | Proteins (most vaccines, toxins) | Polysaccharides, repetitive epitopes (LPS, flagellin) |
| T cell help required | Yes — CD4⁺ Tfh cells | No |
| Germinal centre formation | Yes | No (TI-1) or minimal (TI-2) |
| Class switching | Yes — all isotypes | Limited |
| Affinity maturation | Yes | No |
| Memory formation | Robust, long-lived | Limited |
| Clinical example | Tetanus toxoid, hepatitis B vaccine | Pneumococcal polysaccharide (23vPPV) |
CD4⁺ T Helper Cell Functions
CD4⁺ T helper cells are the orchestrators of adaptive immunity. Upon activation, naïve CD4⁺ T cells differentiate into specialised subsets based on the cytokine environment, each with distinct effector functions relevant to B cell help and broader immune regulation.
Major CD4⁺ T Helper Subsets
| Subset | Key Transcription Factor | Signature Cytokines | Primary Function | B Cell Relevance |
|---|---|---|---|---|
| Th1 | T-bet | IFN-γ, TNF-α, IL-2 | Intracellular pathogens, cell-mediated immunity | Promotes IgG2 (opsonisation, complement fixation) |
| Th2 | GATA-3 | IL-4, IL-5, IL-13 | Helminth defence, allergy | IL-4 drives IgG1 and IgE class switching |
| Th17 | RORγt | IL-17A, IL-17F, IL-22 | Extracellular bacteria, fungi; mucosal defence | Indirect role in mucosal IgA |
| Tfh | Bcl-6 | IL-21, IL-4 | Germinal centre support, B cell help | Primary B cell helper — essential for GC reaction |
| Treg | FoxP3 | IL-10, TGF-β, IL-35 | Immune tolerance, suppression | Regulates GC output, prevents autoantibody production |
T Follicular Helper (Tfh) Cells — The Master B Cell Helpers
Tfh cells are the dominant CD4⁺ subset providing direct help to B cells within germinal centres. They are identified by surface expression of CXCR5 (chemokine receptor directing migration to follicles), PD-1, ICOS, and CD40L, with Bcl-6 as the lineage-defining transcription factor.
Tfh cell functions in germinal centres:
- IL-21 secretion: Major B cell growth and differentiation factor; promotes plasma cell generation and class-switch recombination
- IL-4 secretion: Drives IgG1 and IgE switching
- CD40L expression: Provides survival, proliferation, and differentiation signals to B cells; upregulates AID (activation-induced cytidine deaminase) for somatic hypermutation
- ICOS–ICOSL interaction: Sustains Tfh function within the germinal centre; ICOS deficiency causes a combined immunodeficiency with absent germinal centres
- CXCL13 production: Chemokine that attracts CXCR5⁺ B cells and maintains follicular architecture
Cytokine-Directed Class Switch Recombination
The isotype of antibody produced by B cells is dictated by cytokines from T helper cells during the germinal centre reaction:
Germinal Centre Reaction
The germinal centre (GC) is a specialised microanatomical structure within secondary lymphoid follicles where antigen-activated B cells undergo rapid proliferation, somatic hypermutation (SHM), affinity-based selection, and class-switch recombination (CSR) to generate high-affinity, class-switched antibodies. The GC reaction is the cornerstone of effective humoral immunity and the immunological basis for vaccination.
Germinal Centre Architecture
GCs form in B cell follicles 7–10 days after antigen exposure and consist of two functionally distinct zones:
The Cyclic Re-entry Model
GC B cells cycle between dark and light zones in a process of iterative mutation and selection:
Somatic Hypermutation and Class-Switch Recombination
Both SHM and CSR are catalysed by AID (encoded by AICDA gene):
| Process | Mechanism | Outcome | Clinical Deficiency |
|---|---|---|---|
| Somatic Hypermutation | AID deaminates cytosine → uracil in Ig V-region DNA; error-prone repair introduces point mutations | Altered BCR affinity (increase or decrease); selection retains high-affinity variants | AID deficiency (Hyper-IgM syndrome type 2): absent SHM, impaired CSR, elevated IgM, susceptibility to infections |
| Class-Switch Recombination | AID targets switch (S) regions upstream of constant-region genes; double-strand breaks allow deletion of intervening DNA and recombination to downstream isotype (IgG, IgA, IgE) | Change from IgM/IgD to IgG, IgA, or IgE while retaining the same antigen specificity | UNG deficiency (Hyper-IgM syndrome type 5), NEMO deficiency (X-linked ectodermal dysplasia with immunodeficiency) |
Memory Formation
Immunological memory — the ability to mount faster, stronger, and more specific responses upon re-exposure to a previously encountered antigen — is the fundamental goal of vaccination. Memory is mediated by two major cellular compartments arising from the germinal centre and extrafollicular responses: memory B cells and long-lived plasma cells. Memory T cells provide cellular recall immunity.
Memory B Cell Formation
Memory B cells emerge from germinal centres (GC-derived memory) or from early extrafollicular responses. Key characteristics:
- Surface phenotype: CD27⁺ (in humans), class-switched (IgG⁺, IgA⁺, IgE⁺) or unswitched (IgM⁺IgD⁺CD27⁺ — "IgM memory" from T-independent or marginal zone responses)
- Affinity status: Carry somatically mutated, affinity-matured BCR from GC selection
- Longevity: Can persist for decades; maintenance does not require continuous antigen exposure (debate ongoing; bone marrow niches may play a role)
- Secondary response: Upon re-encounter with antigen, memory B cells rapidly differentiate into antibody-secreting plasma cells or re-enter GCs for further affinity maturation
- Class switching flexibility: Can undergo further class switching upon restimulation (e.g., switch from IgG1 to IgG4 with repeated antigen exposure — relevant in allergen immunotherapy)
Long-Lived Plasma Cells (LLPCs)
LLPCs are terminally differentiated antibody-secreting cells that migrate to survival niches in the bone marrow and constitutively secrete antibody for years to decades without requiring further antigen stimulation.
- Surface phenotype: CD38⁺⁺ CD138⁺ (syndecan-1), CD19⁻/low, CD20⁻, CD27⁺⁺
- Survival signals: BAFF, APRIL (from stromal cells), CXCL12 (from bone marrow niches), IL-6, contact with bone marrow stromal cells
- Antibody secretion rate: Each LLPC secretes ~10,000 antibody molecules per second
- Lifespan: Estimated years to decades; recent evidence from smallpox-vaccinated individuals shows detectable antibodies >50 years post-vaccination
- Niche competition: LLPCs compete for limited bone marrow survival niches; new GC reactions may displace older LLPCs (relevant to repeated vaccination)
Memory T Cell Subsets
Memory T cells provide rapid effector function upon re-encounter with antigen and are categorised by location and function:
| Subset | Location | Surface Markers | Function |
|---|---|---|---|
| Central Memory (Tcm) | Secondary lymphoid organs, blood | CCR7⁺ CD62L⁺ CD45RO⁺ | High proliferative capacity; provide sustained effector supply upon restimulation |
| Effector Memory (Tem) | Peripheral tissues, blood | CCR7⁻ CD62L⁻ CD45RO⁺ | Rapid effector function (cytokine production, cytotoxicity); patrol peripheral tissues |
| Tissue-Resident Memory (Trm) | Non-lymphoid tissues (lung, gut, skin) | CD69⁺ CD103⁺ | First-line defence at barrier sites; do not recirculate; provide immediate local immunity |
| Stem Cell Memory (Tscm) | Blood, lymphoid organs | CD45RA⁺ CD95⁺ CD122⁺ | Self-renewing; longest-lived memory subset; can regenerate all other memory subsets |
Clinical Applications of Memory Biology
Understanding memory formation informs clinical practice in several domains:
- Vaccine design: Adjuvants (e.g., AS01 in Shingrix®, MF59 in Fluad®) enhance GC reactions and memory formation; aluminium salts promote Th2-biased responses with strong antibody but limited T cell memory
- Immunosuppressive therapy: Rituximab (anti-CD20) depletes memory B cells but spares LLPCs (CD20⁻); patients maintain pre-existing antibodies but cannot mount new humoral responses
- Transplant immunology: Donor-specific memory T and B cells mediate accelerated rejection in sensitised recipients; donor-specific antibody (DSA) from LLPCs causes antibody-mediated rejection
- Primary immunodeficiency: Patients with CVID or XLA have impaired memory formation; rely on regular immunoglobulin replacement (Intragam P® 300–600 mg/kg IV every 3–4 weeks, PBS Authority Required)
Pathophysiology of B–T Cell Collaboration Defects
Defects at any step in B–T cell collaboration can result in immunodeficiency, autoimmunity, or lymphoproliferative disorders. Understanding the specific molecular defect informs diagnosis and management.
| Defect | Gene/Protein | Consequence | Inheritance |
|---|---|---|---|
| X-linked agammaglobulinaemia (XLA) | BTK (Bruton tyrosine kinase) | Block in B cell maturation at pre-B stage; absent mature B cells and immunoglobulins | X-linked recessive |
| Hyper-IgM type 1 | CD40LG (CD40 ligand) | Absent T–B cooperation; no class switching; elevated IgM, absent IgG/IgA/IgE | X-linked recessive |
| Hyper-IgM type 2 | AICDA (AID) | Absent SHM and CSR; elevated IgM | Autosomal recessive |
| Common variable immunodeficiency (CVID) | Multiple (ICOS, TACI, BAFF-R, CD19) | Hypogammaglobulinaemia, impaired GC formation, reduced class-switched antibodies | Variable (often polygenic) |
| ICOS deficiency | ICOS | Absent GC formation, CVID phenotype | Autosomal recessive |
| SAP/SH2D1A deficiency (XLP) | SH2D1A (SAP) | Unstable T–B conjugates; absent GC formation; EBV-triggered haemophagocytic lymphohistiocytosis | X-linked recessive |
| Selective IgA deficiency | Unknown (polygenic) | Absent serum IgA (<0.07 g/L); IgG and IgM normal; mucosal infections | Variable |
Investigations
Assessment of B–T cell collaboration and humoral immunity involves a tiered investigation approach available through Australian pathology services.
Special Populations
ATSI Health Considerations
📚 References
- 1. Victora GD, Nussenzweig MC. Germinal centers. Annual Review of Immunology. 2022;40:413–442. doi:10.1146/annurev-immunol-120419-022408
- 2. Crotty S. T follicular helper cell biology: a decade of discovery and diseases. Immunity. 2019;50(5):1132–1148. doi:10.1016/j.immuni.2019.04.011
- 3. Tangye SG, Ma CS, Brink R, Deenick EK. The good, the bad and the ugly — TFH cells in human health and disease. Nature Reviews Immunology. 2013;13(6):412–426. doi:10.1038/nri3447
- 4. Australia Government Department of Health and Aged Care. National Immunisation Program Schedule. Canberra: Australian Government; 2024.
- 5. Australasian Society of Clinical Immunology and Allergy (ASCIA). Primary Immunodeficiency Guidelines. Sydney: ASCIA; 2024. Available from: https://www.allergy.org.au
- 6. Australian Technical Advisory Group on Immunisation (ATAGI). Australian Immunisation Handbook. Canberra: Australian Government Department of Health and Aged Care; 2024. Available from: https://immunisationhandbook.health.gov.au
- 7. Australian Institute of Health and Welfare (AIHW). Aboriginal and Torres Strait Islander Health Performance Framework 2023. Canberra: AIHW; 2023.
- 8. Slifka MK, Antia R, Whitmire JK, Ahmed R. Humoral immunity due to long-lived plasma cells. Immunity. 1998;8(3):363–372. doi:10.1016/S1074-7613(00)80541-5
- 9. Hammarlund E, Lewis MW, Hansen SG, et al. Duration of antiviral immunity after smallpox vaccination. Nature Medicine. 2003;9(9):1131–1137. doi:10.1038/nm917
- 10. Tangye SG, Al-Herz W, Bousfiha A, et al. Human inborn errors of immunity: 2022 update on the classification from the International Union of Immunological Societies Expert Committee. Journal of Clinical Immunology. 2022;42(7):1473–1507. doi:10.1007/s10875-022-01289-3
- 11. Nutt SL, Hodgkin PD, Tarlinton DM, Corcoran LM. The generation of antibody-secreting plasma cells. Nature Reviews Immunology. 2015;15(3):160–171. doi:10.1038/nri3795
- 12. RHDAustralia (Northern Territory Department of Health and Charles Darwin University). Australian Guidelines for the Prevention, Diagnosis and Management of Acute Rheumatic Fever and Rheumatic Heart Disease. 3rd edition. Darwin: RHDAustralia; 2020. [Referenced for ATSI health context and vaccination strategies]