📋 Key Information Summary
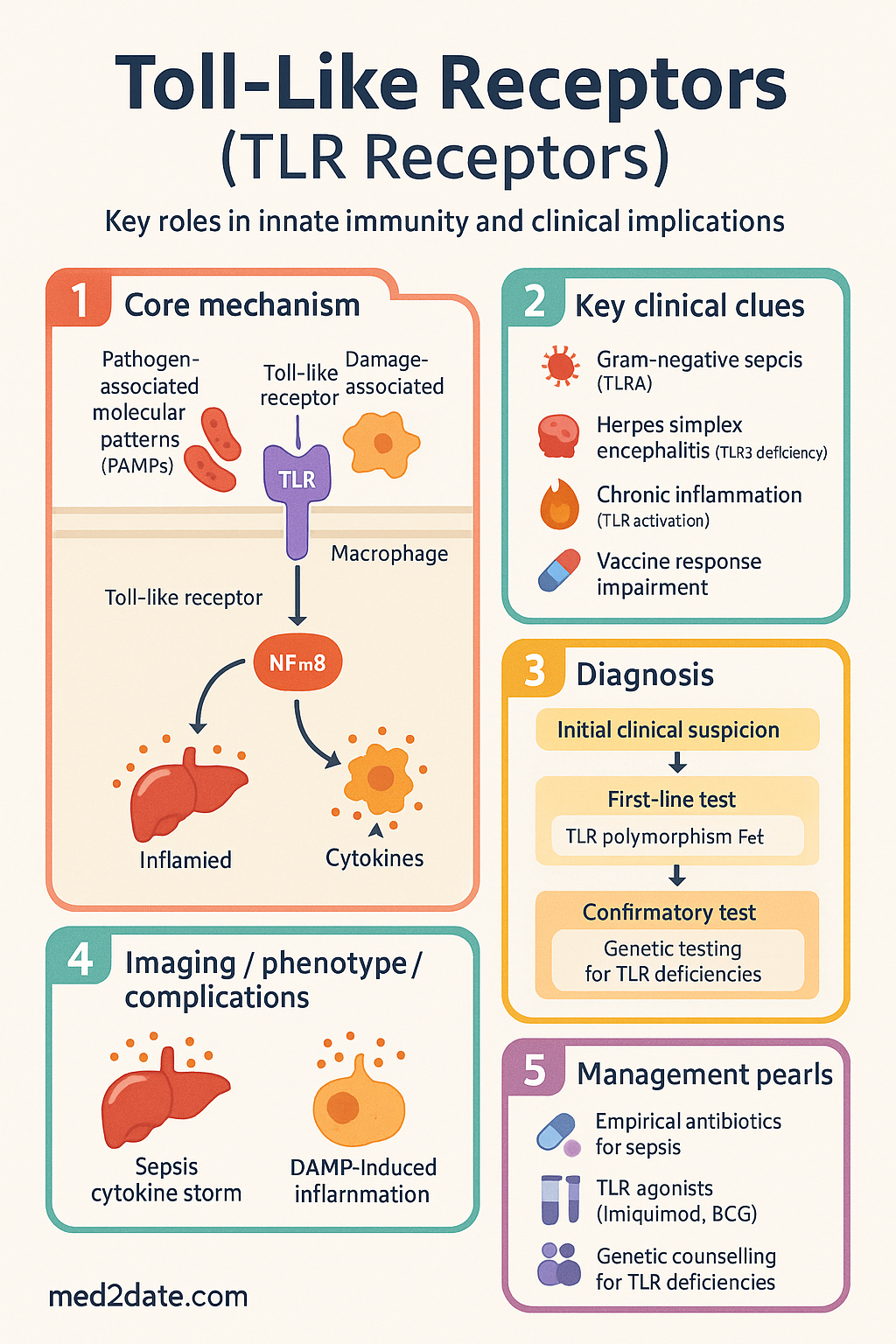
- Toll-like receptors (TLRs) are germline-encoded pattern recognition receptors (PRRs) that detect pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs), forming the first line of innate immune defence.
- Humans express 10 functional TLRs (TLR1–TLR10); TLR1, 2, 4, 5, 6, 10 are surface-localised; TLR3, 7, 8, 9 reside in endosomes and detect nucleic acids.
- TLR4 recognises lipopolysaccharide (LPS) from Gram-negative bacteria — the most clinically studied TLR and central to sepsis pathophysiology in Australian ICUs.
- TLR3 detects double-stranded RNA (dsRNA) and signals exclusively via TRIF, producing robust type I interferon (IFN-α/β) responses critical for antiviral immunity.
- Two major signalling cascades: MyD88-dependent (all TLRs except TLR3) and TRIF/TICAM1-dependent (TLR3 and TLR4), converging on NF-κB, MAPK, and IRF transcription factors.
- Downstream effects include pro-inflammatory cytokine release (TNF-α, IL-1β, IL-6, IL-12), type I interferon production, dendritic cell maturation, and adaptive immune priming.
- Loss-of-function polymorphisms (e.g., TLR4 Asp299Gly, TLR3 deficiency) predispose to severe Gram-negative infections, herpes simplex encephalitis, and impaired vaccine responses.
- Gain-of-function or chronic TLR activation drives pathological inflammation in sepsis, atherosclerosis, inflammatory bowel disease, systemic lupus erythematosus, and rheumatoid arthritis.
- TLR agonists are licensed therapeutics: imiquimod (TLR7, Aldara®) for superficial skin cancers; BCG (multiple TLRs) for bladder cancer; CpG oligonucleotides (TLR9) as vaccine adjuvants.
- TLR-targeted biologics (eritoran — TLR4 antagonist) have not demonstrated mortality benefit in large sepsis trials; current Australian practice relies on source control and empirical antibiotics per eTG.
- Aboriginal and Torres Strait Islander populations carry a higher burden of infections where TLR-mediated immunity is critical; genetic TLR polymorphism studies are ongoing.
- Understanding TLR biology is essential for interpreting vaccine adjuvant mechanisms, primary immunodeficiency workup, and novel immunotherapy strategies in Australian clinical practice.
Introduction & Australian Epidemiological Context
Toll-like receptors (TLRs) are evolutionarily conserved type I transmembrane glycoproteins that function as sentinels of the innate immune system. First characterised through the Drosophila Toll protein in the 1990s and subsequently identified in humans by the laboratory of Bruce Beutler (Nobel Prize in Physiology or Medicine, 2011), TLRs recognise conserved microbial structures — pathogen-associated molecular patterns (PAMPs) — as well as endogenous danger signals released during cellular stress (damage-associated molecular patterns, DAMPs). Activation of TLRs triggers intracellular signalling cascades that culminate in the transcription of pro-inflammatory cytokines, chemokines, type I interferons, and co-stimulatory molecules, thereby shaping both innate and adaptive immune responses.
In Australia, TLR-mediated immunity is clinically relevant across a broad spectrum of infectious and inflammatory conditions. Sepsis accounts for approximately 18 000 hospital separations annually (Australian Institute of Health and Welfare, 2023), with TLR4-mediated recognition of Gram-negative lipopolysaccharide central to the initial cytokine storm. Recurrent herpes simplex encephalitis, a condition linked to TLR3 deficiency, is diagnosed at major tertiary centres including the Royal Children's Hospital Melbourne and Westmead Hospital Sydney. Furthermore, TLR-based therapies — including imiquimod cream (Aldara®) for superficial basal cell carcinoma and BCG intravesical therapy for non-muscle-invasive bladder cancer — are listed on the Pharmaceutical Benefits Scheme (PBS) and prescribed widely across Australian primary care and specialist settings.
This guideline provides a comprehensive overview of TLR types and their ligands, intracellular signalling pathways, downstream immunological effects, and clinical significance relevant to Australian healthcare practitioners, including considerations for Aboriginal and Torres Strait Islander populations and primary immunodeficiency evaluation.
TLR Types & Ligands
Humans express 10 functional Toll-like receptors (TLR1–TLR10). Each TLR recognises specific molecular patterns derived from bacteria, viruses, fungi, or parasites. TLRs are classified by cellular localisation: surface TLRs detect microbial membrane components, while endosomal TLRs detect nucleic acids following phagocytic uptake.
| TLR | Localisation | Primary Ligand (PAMP) | Microbial Source | Key Function |
|---|---|---|---|---|
| TLR1 | Cell surface | Triacyl lipopeptides (with TLR2) | Bacteria, mycobacteria | Heterodimerises with TLR2; senses bacterial lipoproteins |
| TLR2 | Cell surface | Lipoteichoic acid, peptidoglycan, lipoproteins, zymosan | Gram-positive bacteria, fungi, mycobacteria | Broad-spectrum microbial sensor; forms heterodimers with TLR1 or TLR6 |
| TLR3 | Endosome | Double-stranded RNA (dsRNA) | Viruses (e.g., influenza, HSV, West Nile) | Antiviral IFN-α/β production via TRIF |
| TLR4 | Cell surface | Lipopolysaccharide (LPS) | Gram-negative bacteria | Central to sepsis; uses both MyD88 and TRIF pathways; requires MD-2 and CD14 co-receptors |
| TLR5 | Cell surface | Flagellin | Flagellated bacteria (e.g., Salmonella, Pseudomonas) | Mucosal defence in gut and respiratory tract |
| TLR6 | Cell surface | Diacyl lipopeptides (with TLR2) | Mycoplasma, Gram-positive bacteria | Heterodimerises with TLR2; senses mycoplasmal lipoproteins |
| TLR7 | Endosome | Single-stranded RNA (ssRNA) | Viruses (e.g., influenza, HIV, SARS-CoV-2) | Antiviral immunity; therapeutic target (imiquimod) |
| TLR8 | Endosome | Single-stranded RNA (ssRNA) | Viruses, bacteria | Strong TNF-α and IL-12 induction; important in human monocyte activation |
| TLR9 | Endosome | Unmethylated CpG DNA | Bacteria, DNA viruses (HSV, CMV) | IFN-α production; vaccine adjuvant target (CpG ODN) |
| TLR10 | Cell surface | Uncertain (may sense dsRNA or lipopeptides) | Unknown; possibly viral | May function as anti-inflammatory receptor; least characterised |
Endogenous Ligands (DAMPs)
In addition to microbial PAMPs, TLRs recognise endogenous danger signals released during tissue injury, sterile inflammation, and necrotic cell death. These DAMPs include heat shock proteins (HSP60, HSP70), high-mobility group box 1 (HMGB1), fibronectin extra-domain A, hyaluronan fragments, and self-DNA/RNA complexes. Chronic DAMP-driven TLR activation is implicated in atherosclerosis, rheumatoid arthritis, systemic lupus erythematosus, and ischaemia–reperfusion injury — all conditions encountered in Australian clinical practice.
Signalling Pathways
TLR signalling proceeds through two major intracellular pathways: the MyD88-dependent pathway and the TRIF (TICAM1)-dependent pathway. The choice of pathway depends on the TLR involved and determines the transcriptional outcome — predominantly NF-κB-mediated pro-inflammatory cytokine production versus IRF-mediated type I interferon responses.
MyD88-Dependent Pathway
The adaptor protein myeloid differentiation primary response 88 (MyD88) is utilised by all TLRs except TLR3. Upon ligand binding, TLRs recruit MyD88 via homotypic Toll/interleukin-1 receptor (TIR) domain interactions. MyD88 then recruits IL-1 receptor-associated kinases (IRAK4, IRAK1, IRAK2), forming the "Myddosome" complex. IRAK4 phosphorylates IRAK1, which then associates with TNF receptor-associated factor 6 (TRAF6). TRAF6, together with the E2 ubiquitin-conjugating enzyme complex Ubc13/Uev1A, generates K63-linked polyubiquitin chains that activate the TAK1 (TGF-β-activated kinase 1) complex.
TAK1 activates two downstream branches:
- NF-κB pathway: TAK1 phosphorylates the IKK complex (IKKα/IKKβ/NEMO), which degrades IκBα, releasing NF-κB (p50/p65) to translocate to the nucleus and drive transcription of pro-inflammatory cytokines (TNF-α, IL-1β, IL-6, IL-12p40) and co-stimulatory molecules (CD80, CD86).
- MAPK pathway: TAK1 also activates MAP kinase cascades (ERK1/2, JNK, p38), leading to activation of AP-1 transcription factors that synergise with NF-κB for maximal cytokine gene expression.
TRIF-Dependent Pathway
The adaptor TIR-domain-containing adaptor-inducing IFN-β (TRIF, also known as TICAM1) mediates signalling from TLR3 exclusively and from TLR4 in its second (late) signalling phase following endocytosis. TRIF recruits TRAF3, which activates the kinases TBK1 (TANK-binding kinase 1) and IKKε. These kinases phosphorylate interferon regulatory factor 3 (IRF3) and IRF7, promoting their dimerisation, nuclear translocation, and transcription of type I interferons (IFN-α, IFN-β) and interferon-stimulated genes (ISGs).
TRIF also engages TRAF6 and the kinase RIP1 (receptor-interacting protein 1), activating a late-phase NF-κB response independent of MyD88. This dual signalling capacity of TLR4 — early MyD88-dependent and late TRIF-dependent — explains its uniquely broad transcriptional output.
Negative Regulation
Excessive TLR signalling is controlled by multiple negative regulators to prevent immunopathology:
- SIGIRR (TIR8): Decoy TIR-domain receptor that inhibits TLR4 and TLR7/9 signalling in intestinal epithelium.
- A20 (TNFAIP3): Deubiquitinase that removes K63-linked ubiquitin chains from TRAF6 and RIP1, terminating NF-κB signalling.
- SOCS1: Targets MAL/TIRAP for proteasomal degradation, attenuating MyD88-dependent signalling.
- IRAK-M: Inactive IRAK family member that prevents dissociation of IRAK1 from the MyD88 complex.
- MicroRNAs: miR-146a/b downregulates IRAK1 and TRAF6 expression; miR-155 modulates TAB2.
Downstream Effects
TLR activation triggers a coordinated programme of innate and adaptive immune responses. The downstream effects vary by TLR, cell type, and signalling pathway engaged, but collectively serve to eliminate pathogens, recruit immune cells, and initiate adaptive immunity.
Pro-Inflammatory Cytokine & Chemokine Production
NF-κB activation drives transcription of key pro-inflammatory mediators:
| Mediator | Primary Source | Key Functions | Clinical Relevance |
|---|---|---|---|
| TNF-α | Macrophages, monocytes | Fever, acute-phase response, endothelial activation, apoptosis | Central mediator of septic shock; target of anti-TNF biologics in RA |
| IL-1β | Macrophages, dendritic cells | Fever, neutrophil recruitment, Th17 polarisation | Requires NLRP3 inflammasome cleavage; target of anakinra (Kineret®) |
| IL-6 | Macrophages, endothelium | Acute-phase protein induction (CRP, fibrinogen), B-cell differentiation | Elevated in sepsis, RA, Castleman disease; target of tocilizumab |
| IL-12 | Dendritic cells, macrophages | Th1 polarisation, IFN-γ induction in NK and T cells | Critical for intracellular pathogen defence (mycobacteria, Leishmania) |
| IL-10 | Macrophages, Tregs | Anti-inflammatory; limits tissue damage | Regulatory counterbalance; deficiency causes inflammatory bowel disease |
| CXCL8 (IL-8) | Macrophages, epithelium | Neutrophil chemotaxis | Rapid neutrophil recruitment to sites of bacterial infection |
Type I Interferon Production
TLR3, TLR7, TLR8, and TLR9 (endosomal nucleic acid-sensing TLRs) activate IRF3/IRF7 via TRIF or MyD88, driving transcription of IFN-α (13 subtypes) and IFN-β. Type I interferons induce an antiviral state in neighbouring cells by upregulating hundreds of interferon-stimulated genes (ISGs) including Mx proteins, OAS/RNase L, PKR, IFITM proteins, and APOBEC3G. This response is critical for containment of viral infections including influenza, SARS-CoV-2, and herpes simplex virus.
Dendritic Cell Maturation & Adaptive Immune Priming
TLR activation in dendritic cells (DCs) triggers a maturation programme essential for bridging innate and adaptive immunity:
- Upregulation of MHC class II: Enhanced antigen presentation to CD4+ T cells.
- Expression of co-stimulatory molecules: CD80 (B7-1), CD86 (B7-2), and CD40 provide Signal 2 for T-cell activation, preventing anergy.
- Cytokine-directed T-cell polarisation: IL-12 drives Th1; IL-6 + TGF-β drives Th17; IFN-α enhances cytotoxic T lymphocyte (CTL) responses.
- Lymph node migration: Upregulation of CCR7 directs DCs to T-cell zones of draining lymph nodes.
Inflammasome Cross-Talk
TLR signalling provides "Signal 1" (priming) for NLRP3 inflammasome activation by upregulating pro-IL-1β and NLRP3 expression. A second signal (e.g., ATP, crystalline urate, bacterial toxins) then triggers inflammasome assembly, caspase-1 activation, and mature IL-1β/IL-18 secretion. This two-step model is clinically relevant in gout, pseudogout, and silicosis — conditions managed by Australian rheumatologists.
Clinical Significance
TLR biology has direct relevance to multiple areas of Australian clinical medicine, including infectious diseases, immunodeficiency, rheumatology, oncology, and vaccinology.
Sepsis & TLR4
TLR4-mediated recognition of LPS is the initiating event in Gram-negative sepsis. Activation triggers massive TNF-α, IL-1β, and IL-6 release, leading to vasodilation, capillary leak, disseminated intravascular coagulation (DIC), and multi-organ failure. TLR4 also senses DAMPs (HMGB1, HSPs) during later phases, perpetuating inflammation even after pathogen clearance. TLR4 antagonists (eritoran, TAK-242) showed promise in preclinical models but failed to demonstrate mortality benefit in large randomised controlled trials (ACCESS trial, 2013). Current Australian management of sepsis prioritises source control, empirical antibiotics, and haemodynamic resuscitation per the Surviving Sepsis Campaign guidelines, adopted in Australian ICUs.
Primary Immunodeficiency & TLR Defects
Genetic defects in TLR signalling components cause specific primary immunodeficiency syndromes:
| Defect | Gene | TLR Affected | Clinical Phenotype | Australian Relevance |
|---|---|---|---|---|
| MyD88 deficiency | MYD88 | All except TLR3 | Recurrent pyogenic bacterial infections (S. pneumoniae, S. aureus) in childhood | Diagnosed at paediatric immunology centres (RCH Melbourne, Sydney Children's) |
| IRAK-4 deficiency | IRAK4 | All except TLR3 | Invasive pneumococcal disease, other pyogenic infections; improves with age | Included in expanded newborn screening discussions |
| TLR3 deficiency | TLR3 | TLR3 | Herpes simplex encephalitis (HSE) in otherwise healthy children/adults | Consider in recurrent or atypical HSE presentations |
| UNC93B1 deficiency | UNC93B1 | TLR3, 7, 8, 9 | Herpes simplex encephalitis | Functional TLR3 assay available at reference laboratories |
TLR-Based Therapeutics
Several TLR-targeted agents are available in Australia:
Autoimmune & Inflammatory Disease
Chronic or inappropriate TLR activation contributes to the pathogenesis of several autoimmune conditions managed in Australian specialist practice:
- Systemic lupus erythematosus (SLE): Immune complexes containing self-DNA and self-RNA activate TLR9 and TLR7, respectively, in plasmacytoid dendritic cells, driving IFN-α overproduction — the "interferon signature" characteristic of SLE. Hydroxychloroquine, a mainstay of SLE therapy in Australia, inhibits endosomal TLR activation by raising endosomal pH.
- Rheumatoid arthritis (RA): TLR2 and TLR4 are upregulated on synovial macrophages; their activation by DAMPs (fibronectin fragments, HSPs, HMGB1) sustains synovial inflammation and joint destruction.
- Inflammatory bowel disease (IBD): TLR signalling in intestinal epithelial cells maintains barrier integrity; dysregulated TLR2/4 responses contribute to Crohn's disease pathogenesis. SIGIRR (TIR8) polymorphisms are associated with IBD susceptibility.
- Atherosclerosis: TLR2 and TLR4 on macrophages recognise oxidised LDL and DAMPs within atherosclerotic plaques, promoting foam cell formation and plaque instability.
Vaccine Adjuvants
TLR agonists are increasingly used as vaccine adjuvants to enhance immunogenicity:
- AS01 (shingles vaccine — Shingrix®): Contains MPL (TLR4 agonist) plus QS-21 saponin; achieves >90% efficacy against herpes zoster even in immunocompromised patients. Available on NIP for adults ≥65 years and immunocompromised patients ≥18 years in Australia.
- AS04 (HPV vaccine — Cervarix®): MPL adsorbed to aluminium salt (TLR4 agonist); induces stronger and more durable antibody responses than aluminium alone.
- CpG 1018 (Heplisav-B®): TLR9 agonist adjuvant for hepatitis B vaccine; 2-dose schedule with higher seroprotection rates. Not yet PBS-listed in Australia but available privately.
TLR Polymorphisms & Susceptibility
Common single nucleotide polymorphisms (SNPs) in TLR genes modulate infection susceptibility and disease severity:
| Polymorphism | Gene | Effect | Clinical Association |
|---|---|---|---|
| Asp299Gly | TLR4 | Reduced LPS responsiveness | Increased Gram-negative infection risk; reduced atherosclerosis |
| Thr399Ile | TLR4 | Reduced LPS responsiveness | Haplotype-dependent; associated with malaria resistance |
| Arg753Gln | TLR2 | Impaired lipopeptide recognition | Increased susceptibility to tuberculosis, leprosy |
| P554S | TLR3 | Loss of function | Herpes simplex encephalitis in children |
| −1237C | TLR9 | Increased transcriptional activity | Associated with SLE susceptibility in some populations |
Pathophysiology
The pathophysiological role of TLRs extends from protective antimicrobial immunity to pathological inflammation in autoimmunity, chronic infection, and malignancy. Understanding the balance between protective TLR activation and harmful dysregulation is central to clinical application.
Protective Immunity
Under normal physiological conditions, TLR activation provides rapid, localised antimicrobial defence:
- Epithelial barrier defence: TLR2, TLR4, and TLR5 on mucosal epithelial cells (gut, respiratory tract, urinary tract) detect commensal-derived and pathogenic signals, maintaining barrier homeostasis through antimicrobial peptide (defensin, cathelicidin) production and tight junction regulation.
- Phagocyte activation: Macrophages and neutrophils upregulate reactive oxygen species (ROS) production, phagolysosomal killing, and neutrophil extracellular trap (NET) formation following TLR engagement.
- Antiviral state: Endosomal TLR3/7/8/9-mediated IFN-α/β production induces an antiviral state in uninfected cells, limiting viral spread.
Pathological Hyperactivation — Sepsis Paradigm
Excessive TLR signalling during overwhelming infection leads to the systemic inflammatory response syndrome (SIRS) characteristic of sepsis:
- Massive TNF-α and IL-1β release causes systemic vasodilation, capillary leak, and hypotension.
- IL-6 drives hepatic acute-phase response (CRP, ferritin, fibrinogen) and contributes to DIC via tissue factor upregulation.
- Endothelial activation and coagulation cascade activation lead to microvascular thrombosis and organ ischaemia.
- Late-phase DAMP signalling (HMGB1 → TLR4) perpetuates inflammation even after successful antimicrobial therapy, contributing to immunoparalysis.
Sterile Inflammation & Autoimmunity
In the absence of infection, TLR activation by endogenous DAMPs drives pathological sterile inflammation:
- SLE: Self-nucleic acids in immune complexes activate TLR7 (ssRNA) and TLR9 (DNA) on plasmacytoid DCs, producing pathological IFN-α. This drives B-cell activation, autoantibody production (anti-dsDNA, anti-Smith), and immune complex deposition.
- Gout: Monosodium urate (MSU) crystals provide Signal 2 for NLRP3 inflammasome activation following TLR priming (Signal 1) by gut-derived LPS or endogenous DAMPs.
- Atherosclerosis: Oxidised phospholipids and HMGB1 in atherosclerotic plaques activate TLR2/4 on macrophages, perpetuating inflammation and promoting plaque rupture.
Tumour Immune Evasion
Tumours exploit TLR signalling for immune evasion and growth promotion. Tumour-derived exosomes carrying HSP70 and HMGB1 activate TLR2/4 on myeloid-derived suppressor cells (MDSCs), promoting immunosuppressive microenvironments. Conversely, therapeutic TLR activation (imiquimod, BCG) can overcome immune evasion by restoring anti-tumour immunity.
Investigations
TLR pathway assessment is primarily indicated in the evaluation of primary immunodeficiency, recurrent or atypical infections, and research settings. Clinical TLR functional testing is available through specialised immunology laboratories in Australia.
Monitoring
Monitoring in TLR-related conditions focuses on disease activity in autoimmune conditions driven by chronic TLR activation, response to TLR-based therapeutics, and immunodeficiency surveillance.
| Condition | Monitoring Parameters | Frequency |
|---|---|---|
| SLE (TLR7/9-driven IFN-α) | Anti-dsDNA titre, complement (C3/C4), urinalysis, CRP/ESR, SLEDAI-2K score, IFN-α gene signature | Every 1–3 months if active; 6-monthly if stable |
| Sepsis (TLR4-mediated) | Lactate, CRP, procalcitonin, organ function (creatinine, bilirubin, platelets), SOFA score | Continuous haemodynamic; labs every 6–12 hours in ICU |
| Imiquimod therapy (TLR7 agonist) | Local skin reaction (erythema, erosion), treatment response (clinical ± dermoscopy), systemic side effects (flu-like symptoms) | 4-weekly during treatment; post-treatment review at 3 months |
| BCG intravesical (TLR2/4 agonist) | Cystoscopy with biopsies, urine cytology, adverse events (cystitis, haematuria, BCG sepsis) | 3-monthly cystoscopy for 2 years, then 6-monthly to 5 years |
| Primary immunodeficiency (MyD88/IRAK-4) | Infection frequency, immunoglobulin levels (IgG, IgA, IgM), vaccine responses (pneumococcal serology), growth and development | 3–6-monthly with clinical immunologist |
Special Populations
Aboriginal and Torres Strait Islander Health Considerations
Aboriginal and Torres Strait Islander peoples experience a disproportionate burden of infectious and inflammatory diseases where TLR-mediated immunity plays a central role. Understanding these disparities is essential for culturally safe and clinically effective healthcare.
📚 References
- 1. Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nature Immunology. 2010;11(5):373–384.
- 2. O'Neill LAJ, Golenbock D, Bowie AG. The history of Toll-like receptors — redefining innate immunity. Nature Reviews Immunology. 2013;13(6):453–460.
- 3. Takeuchi O, Akira S. Pattern recognition receptors and inflammation. Cell. 2010;140(6):805–820.
- 4. Opal SM, Laterre PF, François B, et al. Effect of eritoran, an antagonist of MD2-TLR4, on mortality in patients with severe sepsis: the ACCESS randomized controlled trial. JAMA. 2013;309(11):1154–1162.
- 5. Picard C, Casanova JL, Puel A. Infectious diseases in patients with IRAK-4, MyD88, NEMO, or IκBα deficiency. Clinical Microbiology Reviews. 2011;24(3):490–497.
- 6. Zhang SY, Jouanguy E, Ugolini S, et al. TLR3 deficiency in patients with herpes simplex encephalitis. Science. 2007;317(5844):1522–1527.
- 7. Australian Institute of Health and Welfare (AIHW). Sepsis in Australia. Cat. no. PHE 317. Canberra: AIHW; 2023.
- 8. Australian Commission on Safety and Quality in Health Care (ACSQHC). Sepsis Clinical Care Standard. Sydney: ACSQHC; 2022.
- 9. National Health and Medical Research Council (NHMRC). Values and Ethics: Guidelines for Ethical Conduct in Aboriginal and Torres Strait Islander Health Research. Canberra: NHMRC; 2018.
- 10. Casanova JL, Abel L, Quintana-Murci L. Human TLRs and IL-1Rs in host defense: natural insights from evolutionary, epidemiological, and clinical genetics. Annual Review of Immunology. 2011;29:447–491.
- 11. Bryant CE, Monie TP. Mice, men and the relatives: cross-species studies underpin innate immunity. Open Biology. 2012;2(4):120015.
- 12. Lancet Infectious Diseases Commission. Vaccine adjuvants: putting innate immunity to work. Lancet Infectious Diseases. 2020;20(10):e250–e261.
- 13. AIHW. Aboriginal and Torres Strait Islander Health Performance Framework 2020 summary report. Canberra: AIHW; 2020.
- 14. ANZDATA Registry. 45th Annual Report 2022 (Data to 2021). Adelaide: Australia and New Zealand Dialysis and Transplant Registry; 2022.
- 15. Rischmueller M, Tieu J, Lester S. Primary Sjögren's syndrome. Best Practice & Research Clinical Rheumatology. 2016;30(1):189–220.