📋 Key Information Summary
- Dendritic cells (DCs) are the most potent professional antigen-presenting cells (APCs), bridging innate and adaptive immunity through antigen capture, processing, and presentation to naïve T cells.
- Two principal subtypes: Plasmacytoid DCs (pDCs) produce massive type I interferon (IFN-α/β) in response to viral nucleic acids via TLR7/9; Myeloid/conventional DCs (cDCs) are the primary initiators of T-cell priming via MHC-I and MHC-II presentation.
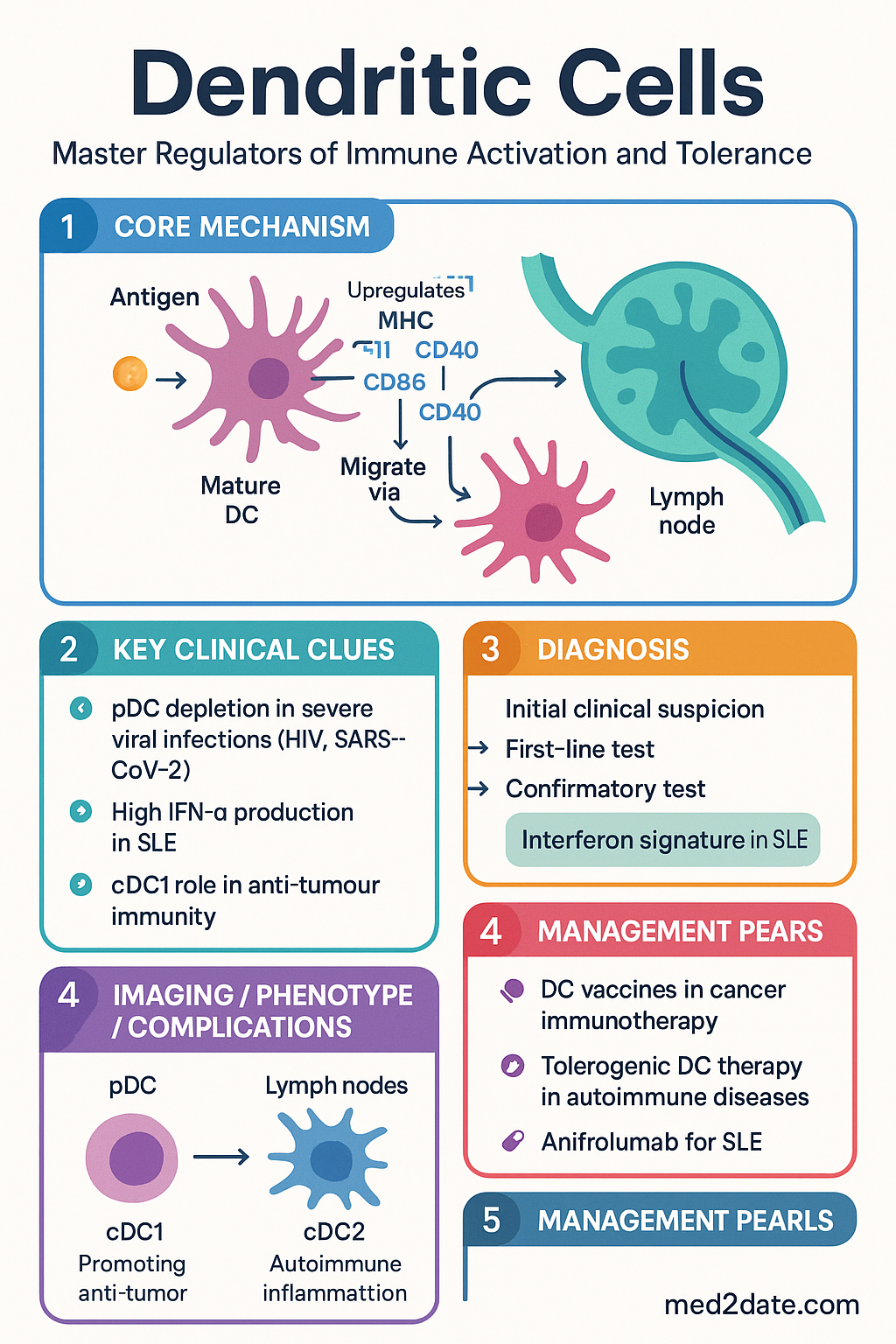
- Maturation is the critical switch — immature DCs capture antigen efficiently but express low MHC and co-stimulatory molecules; upon activation by PAMPs or DAMPs, mature DCs upregulate CD80, CD86, CD40, and CCR7 to migrate to lymph nodes.
- Migration to T-cell zones of secondary lymphoid organs is governed by a chemokine gradient shift: from CCR1/CCR5/CCR6 (inflammatory sites) to CCR7 (lymph node T-cell zones) upon maturation.
- Antigen presentation occurs via two major pathways: MHC class I (endogenous/cross-presented antigens → CD8⁺ T cells) and MHC class II (exogenous antigens → CD4⁺ T cells), with cross-presentation being a unique DC capability critical for anti-tumour and anti-viral immunity.
- Tolerance vs immunity — the outcome depends on DC activation state: immature/tolerogenic DCs promote T-cell anergy, deletion, or Treg induction (peripheral tolerance), while mature DCs drive effector T-cell differentiation (immunity).
- Clinical relevance in Australia: DC biology underpins cancer immunotherapy (DC vaccines approved in trials at Peter MacCallum, Westmead), autoimmune disease pathogenesis (SLE, type 1 diabetes), transplant tolerance strategies, and adjuvant design for national vaccination programmes.
- cDC1 (BATF3⁺) are specialised cross-presenting DCs essential for CD8⁺ T-cell responses against tumours and intracellular pathogens; cDC2 (IRF4⁺) primarily drive CD4⁺ T-cell responses including Th2 and Th17.
- pDC depletion is a hallmark of severe viral infections (HIV, SARS-CoV-2) and correlates with impaired IFN responses; Australian research from the Kirby Institute and WEHI has contributed significantly to understanding pDC dysfunction in chronic viral disease.
- Tolerogenic DC therapy is an emerging frontier: ex vivo-generated tolerogenic DCs pulsed with autoantigens are under clinical investigation for type 1 diabetes, rheumatoid arthritis, and multiple sclerosis at Australian research centres.
- Pattern recognition receptors on DCs (TLRs, CLRs, RLRs, NLRs) integrate diverse danger signals — this receptor repertoire determines whether an immune response favours tolerance or immunity.
- DC dysfunction underlies immunodeficiency (IRF8, GATA2 mutations), autoimmunity (impaired tolerogenic function), and chronic infection (exhaustion phenotypes), with diagnostic and therapeutic implications for Australian haematology and immunology services.
Introduction & Australian Context
Dendritic cells (DCs) are the most potent professional antigen-presenting cells in the human immune system, functioning as sentinels that bridge innate and adaptive immunity. First described by Ralph Steinman and Zanvil Cohn in 1973 at the Rockefeller University — work that earned Steinman the 2011 Nobel Prize in Physiology or Medicine — DCs are now recognised as the master regulators of immune activation and tolerance.
DCs reside in peripheral tissues (skin, mucosal surfaces, solid organs) in an immature state, continuously sampling the antigenic environment through macropinocytosis, receptor-mediated endocytosis, and phagocytosis. Upon encountering pathogen-associated molecular patterns (PAMPs) or damage-associated molecular patterns (DAMPs), DCs undergo a dramatic phenotypic and functional transformation known as maturation, which includes upregulation of MHC molecules, co-stimulatory ligands (CD80, CD86, CD40), and the lymph node–homing chemokine receptor CCR7.
In the Australian clinical context, DC biology is relevant across multiple disciplines. At the Peter MacCallum Cancer Centre (Melbourne) and Westmead Hospital (Sydney), DC-based immunotherapies are in active clinical trials for melanoma, prostate cancer, and glioblastoma. The Garvan Institute of Medical Research and WEHI (Walter and Eliza Hall Institute) have made internationally recognised contributions to understanding DC subsets, cross-presentation, and tolerogenic DC programming. DC dysfunction is implicated in the pathogenesis of systemic lupus erythematosus (pDC-derived IFN-α), type 1 diabetes, and transplant rejection — conditions managed daily in Australian tertiary centres.
This guideline provides a comprehensive overview of DC biology with emphasis on clinical translation: DC subtypes and their distinct roles, the maturation and migration programme, antigen presentation pathways, and the critical balance between tolerance and immunity.
Dendritic Cell Subtypes
Human dendritic cells are heterogeneous, comprising multiple subsets with distinct developmental origins, surface phenotypes, tissue distributions, and functional specialisations. The two principal lineages — plasmacytoid DCs (pDCs) and myeloid/conventional DCs (cDCs) — serve complementary but non-overlapping roles in immune surveillance.
Plasmacytoid Dendritic Cells (pDCs)
pDCs are specialised sentinels of the innate antiviral immune response. Named for their morphological resemblance to plasma cells, pDCs circulate in the blood and home to lymphoid tissues, where they are strategically positioned to detect viral nucleic acids.
| Feature | Detail |
|---|---|
| Surface markers | CD123 (IL-3Rα), CD303 (BDCA-2/CLEC4C), CD304 (Neuropilin-1), CD45RA, ILT7 |
| Key transcription factors | E2-2 (TCF4), SPIB, IRF7, IRF8 |
| Pattern recognition | TLR7 (ssRNA), TLR9 (CpG DNA) — endosomal localisation restricts detection to nucleic acids |
| Signature function | Massive type I interferon production (IFN-α, IFN-β) — up to 1,000× more than other cell types |
| Antigen presentation | Weaker than cDCs; can present via MHC-I and MHC-II but primarily function as cytokine producers rather than T-cell primers |
| Tissue distribution | Blood, T-cell zones of lymph nodes, mucosal-associated lymphoid tissue (MALT), bone marrow |
| Clinical significance | IFN-α production drives lupus pathogenesis (SLE); pDC depletion in HIV, SARS-CoV-2 correlates with disease severity |
Myeloid / Conventional Dendritic Cells (cDCs)
cDCs are the classical antigen-presenting DCs and are further subdivided into two major subsets based on ontogeny, transcription factor dependence, and functional specialisation.
| Feature | cDC1 | cDC2 |
|---|---|---|
| Surface markers | CD141 (BDCA-3/CLEC9A), XCR1, CD11c | CD1c (BDCA-1), CD11c, CD172a (SIRPα) |
| Key transcription factors | BATF3, IRF8, ID2 | IRF4, ZEB2, KLF4 |
| Primary function | Cross-presentation via MHC-I → CD8⁺ cytotoxic T-cell priming | MHC-II presentation → CD4⁺ T-cell priming (Th1, Th2, Th17) |
| Pattern recognition | TLR3 (dsRNA), TLR8 (ssRNA), CLEC9A (dead cell–associated actin) | TLR1/2/4/5/6/8, DC-SIGN, Dectin-1, Mannose receptor |
| Cytokine production | IL-12p70, IFN-λ | IL-1β, IL-6, IL-12, IL-23, TNF-α |
| Clinical role | Anti-tumour immunity, anti-viral CD8⁺ responses | Anti-bacterial/fungal immunity, allergic sensitisation, Th17 responses |
Other DC-Related Populations
| Population | Characteristics | Clinical Relevance |
|---|---|---|
| Langerhans cells (LCs) | Epidermal DCs; CD1a⁺, Langerin (CD207)⁺, Birbeck granules; self-renewing tissue-resident population | Langerhans cell histiocytosis (LCH) — BRAF V600E mutation common; managed at Australian paediatric oncology centres |
| Monocyte-derived DCs (moDCs) | Differentiate from CD14⁺ monocytes in inflammation; CD1a⁺/CD14⁺; short-lived; major DC population at inflammatory sites | Used ex vivo to generate DC vaccines for cancer immunotherapy |
| Inflammatory DCs (Tip-DCs) | TNF/iNOS-producing DCs; CD11c⁺, CD11b⁺; recruited to sites of infection | Contribute to tissue damage in severe infections and autoimmunity |
| AS-DC (AXL⁺/SIGLEC6⁺) | Recently identified transitional population; pDC-like but with cDC antigen-presenting capacity | Implicated in SLE pathogenesis alongside classical pDCs |
Maturation & Migration
The transition from an immature, antigen-capturing sentinel to a mature, T cell–activating antigen-presenting cell is the defining functional programme of dendritic cells. This process — termed maturation — is accompanied by a coordinated migration from peripheral tissues to the T-cell zones of draining lymph nodes.
Signals That Drive Maturation
| Signal Category | Examples | Receptor / Pathway |
|---|---|---|
| PAMPs | LPS, flagellin, dsRNA, CpG DNA, viral ssRNA, β-glucan | TLR1–10, RIG-I/MDA5, CLRs (Dectin-1/2, Mincle), cGAS-STING |
| DAMPs | HMGB1, ATP, uric acid crystals, heat-shock proteins, self-DNA/RNA | RAGE, P2X7, NLRP3 inflammasome, TLR4 |
| Inflammatory cytokines | TNF-α, IL-1β, IL-6, IFN-γ, type I IFNs | Respective cytokine receptors; NF-κB and JAK-STAT signalling |
| T-cell signals | CD40L (CD154) on activated T cells | CD40 on DCs — "licence" signal for full maturation and sustained IL-12 production |
| Complement | C3a, C5a | C3aR, C5aR on DCs; enhances maturation and migration |
Phenotypic and Functional Changes During Maturation
The Migration Programme
DC migration is a chemokine-directed, multi-step process that repositions antigen-loaded DCs from peripheral tissues to the T-cell zones of draining lymph nodes:
- Tissue exit: Maturation triggers downregulation of tissue-retention chemokine receptors (CCR1, CCR5, CCR6) and upregulation of CCR7. CCR7 responds to CCL19 (ELC) and CCL21 (SLC) produced by lymphatic endothelial cells and lymph node stroma.
- Afferent lymphatic transit: DCs enter afferent lymphatics, traversing the vessel wall via afferent lymphatic endothelial cells. DCs acquire a characteristic elongated morphology to navigate narrow lymphatic capillaries.
- Lymph node entry: DCs arrive at the subcapsular sinus and migrate into the T-cell zone (paracortex) guided by CCL21 gradients.
- T-cell scanning: In the paracortex, mature DCs extend dendrites and form immunological synapses with naïve T cells, scanning up to 500–5,000 T cells per hour for cognate TCR–MHC–peptide interactions.
- Transcytosis route: In mucosal tissues (gut, respiratory tract), DCs can also extend processes across epithelial barriers to sample luminal antigens without leaving the tissue, or transport antigens to lymph nodes via transcytosis through goblet cells.
Antigen Presentation
Antigen presentation is the cardinal function of dendritic cells and the molecular basis for their role as the initiators of adaptive immunity. DCs present processed peptide fragments on major histocompatibility complex (MHC) molecules to T-cell receptors (TCRs), providing three signals necessary for T-cell activation: (1) antigen recognition (signal 1), (2) co-stimulation (signal 2), and (3) polarising cytokines (signal 3).
MHC Class II Pathway (Exogenous Antigens)
The classical pathway for presenting exogenous antigens to CD4⁺ helper T cells:
- Antigen uptake: Receptor-mediated endocytosis (via CLRs, FcγR, complement receptors), macropinocytosis, or phagocytosis.
- Endosomal processing: Antigens are degraded by cathepsins (S, L, B, D) in progressively acidified endosomal compartments (early endosome → late endosome → lysosome) into 13–25 amino acid peptides.
- MHC-II loading: Newly synthesised MHC-II α/β heterodimers associate with the invariant chain (Ii, CD74) in the ER, which blocks premature peptide binding and directs MHC-II to the endosomal MIIC compartment. HLA-DM catalyses removal of the CLIP fragment and loading of high-affinity peptide.
- Surface expression: Peptide–MHC-II complexes are transported to the cell surface for presentation to CD4⁺ T cells.
MHC Class I Pathway (Endogenous Antigens)
The classical pathway for presenting intracellular antigens to CD8⁺ cytotoxic T cells:
- Cytosolic degradation: Endogenous proteins (including viral proteins in infected cells) are ubiquitinated and degraded by the proteasome (immunoproteasome in activated DCs) into 8–11 amino acid peptides.
- TAP transport: Peptides are translocated into the ER lumen by TAP1/TAP2 (transporter associated with antigen processing).
- MHC-I loading: Peptides are loaded onto MHC-I heavy chain/β2-microglobulin complexes with the assistance of the peptide-loading complex (tapasin, ERp57, calreticulin).
- Surface expression: Stable peptide–MHC-I complexes traffic to the cell surface for CD8⁺ T-cell recognition.
Cross-Presentation — The Unique DC Capability
| Cross-Presentation Pathway | Mechanism | Key Features |
|---|---|---|
| Cytosolic pathway | Exogenous antigens escape from endosomes/phagosomes into the cytosol → proteasomal degradation → TAP-dependent ER loading onto MHC-I | Dependent on Sec61 translocon, ERAD machinery; dominant pathway in cDC1 |
| Vacuolar pathway | Antigens are degraded by cathepsins within the endosome/phagosome → peptides loaded onto recycling MHC-I within the same compartment | TAP-independent, proteasome-independent; faster but less selective |
The Three-Signal Model of T-Cell Activation
Tolerance vs Immunity
The decision between immune tolerance and immunity is not determined by the antigen itself but by the context in which the antigen is presented — and dendritic cells are the arbiters of this decision. Immature and semi-mature DCs presenting self-antigens without inflammatory co-signals drive tolerance, while fully mature DCs presenting pathogen-derived antigens with co-stimulation drive immunity.
Mechanisms of DC-Mediated Tolerance
| Mechanism | Description | Clinical Correlate |
|---|---|---|
| Anergy induction | Immature DCs present antigen (signal 1) without co-stimulation (signal 2). T cells recognise antigen but receive no activation signal, resulting in long-lasting functional unresponsiveness (anergy). | Peripheral tolerance to self-antigens; basis for tolerogenic DC therapy |
| Deletion (apoptosis) | DCs presenting antigen in tolerogenic conditions can induce activation-induced cell death (AICD) via Fas/FasL or TRAIL pathways. | Elimination of autoreactive T cells escaping thymic negative selection |
| Treg induction | Semi-mature DCs producing TGF-β, IL-10, retinoic acid, and expressing low co-stimulatory molecules convert naïve CD4⁺ T cells into FOXP3⁺ regulatory T cells (Tregs). | Gut tolerance (gDCs + retinoic acid → iTregs); transplant tolerance; allergy prevention |
| IDO-mediated suppression | Tolerogenic DCs express indoleamine 2,3-dioxygenase (IDO), which depletes local tryptophan and generates immunosuppressive kynurenines, inhibiting T-cell proliferation. | Maternal–fetal tolerance; tumour immune evasion; transplant acceptance |
| Peripheral tissue-resident DCs | Tissue-resident DCs continuously sample self-antigens and migrate to lymph nodes in a steady-state (non-inflamed) manner, presenting self-peptides in a tolerogenic context. | Steady-state DC migration maintains peripheral tolerance to tissue-restricted antigens |
Tolerogenic DCs — Therapeutic Applications
Ex vivo generation of tolerogenic DCs is a rapidly advancing field with active Australian research participation. These cells are generated by exposing monocyte-derived DCs to immunosuppressive agents or cytokines before pulsing with disease-relevant autoantigens.
When Tolerance Fails — DCs in Autoimmunity
- Systemic lupus erythematosus (SLE): pDC activation by immune complex–nucleic acid → IFN-α storm → B-cell activation → autoantibody production. Anifrolumab (anti-IFNAR) is PBS Authority Required.
- Type 1 diabetes: Failure of pancreatic DC tolerance → autoreactive CD8⁺ T-cell destruction of β-cells. Islet antigen–loaded tolerogenic DCs under investigation.
- Rheumatoid arthritis: Synovial DCs present citrullinated peptides in an immunogenic context → Th17/Th1 activation → joint inflammation.
- Multiple sclerosis: CNS-resident DCs presenting myelin antigens in the context of inflammation → demyelinating T-cell responses.
DCs in Immunity — Initiating Protective Responses
When fully activated by PAMPs and inflammatory signals, mature DCs become the most potent stimulators of naïve T-cell immunity:
pDC-derived IFN-α establishes the antiviral state. cDC1 cross-present viral antigens on MHC-I to prime cytotoxic CD8⁺ T cells. cDC2 present on MHC-II to generate CD4⁺ Th1 help. This three-arm response is the basis for effective viral clearance and underpins vaccine design for influenza, SARS-CoV-2, and HPV programmes funded under the National Immunisation Programme.
cDC1 cross-presentation of tumour-associated antigens on MHC-I is essential for anti-tumour CD8⁺ T-cell priming. Tumour-derived factors (VEGF, IL-10, PGE2, lactic acid) impair DC maturation and function — a key immune evasion mechanism. DC vaccines (sipuleucel-T for prostate cancer; Australian trials in melanoma) and STING agonists aim to overcome tumour-mediated DC suppression.
Aboriginal and Torres Strait Islander Health Considerations
📚 References
- 1. Steinman RM, Cohn ZA. Identification of a novel cell type in peripheral lymphoid organs of mice. I. Morphology, quantitation, tissue distribution. J Exp Med. 1973;137(5):1142–1162.
- 2. Merad M, Sathe P, Helft J, Miller J, Mortha A. The dendritic cell lineage: ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Annu Rev Immunol. 2013;31:563–604.
- 3. Collin M, Bigley V. Human dendritic cell subsets: an update. Immunology. 2018;154(1):3–20.
- 4. Cella M, Jarrossay D, Facchetti F, et al. Plasmacytoid monocytes migrate to inflamed lymph nodes and produce large amounts of type I interferon. Nat Med. 1999;5(8):919–923.
- 5. Hildner K, Edelson BT, Purtha WE, et al. Batf3 deficiency reveals a critical role for CD8α⁺ dendritic cells in cytotoxic T cell immunity. Science. 2008;322(5904):1097–1100.
- 6. Joffre OP, Segura E, Savina A, Amigorena S. Cross-presentation by dendritic cells. Nat Rev Immunol. 2012;12(8):557–569.
- 7. Wculek SK, Cueto FJ, Mujal AM, et al. Dendritic cells in cancer immunology and immunotherapy. Nat Rev Immunol. 2020;20(1):7–24.
- 8. Yogev N, Frommer F, Lukas D, et al. Dendritic cells ameliorate autoimmunity in the CNS by controlling the homeostasis of PD-1 receptor⁺ regulatory T cells. Immunity. 2012;37(2):264–275.
- 9. Cauwels A, Tavernier J. Tolerizing strategies for the treatment of autoimmune diseases: from ex vivo to in vivo strategies. Front Immunol. 2020;11:534.
- 10. Navegantes KC, de Souza Gomes R, Pereira PAT, et al. Immune modulation of some autoimmune diseases: the critical role of macrophages and neutrophils in the innate and adaptive immunity. J Transl Med. 2017;15(1):36.
- 11. Guilliams M, Ginhoux F, Jakubzick C, et al. Dendritic cells, monocytes and macrophages: a unified nomenclature based on ontogeny. Nat Rev Immunol. 2014;14(8):571–578.
- 12. Australian Institute of Health and Welfare (AIHW). Aboriginal and Torres Strait Islander Health Performance Framework 2020 summary report. Canberra: AIHW; 2020.
- 13. RHDAustralia (ARF/RHD writing group). The 2020 Australian guideline for prevention, diagnosis and management of acute rheumatic fever and rheumatic heart disease. 3rd edn. Darwin: Menzies School of Health Research; 2020.
- 14. National Health and Medical Research Council (NHMRC). National Immunisation Program Schedule. Canberra: Australian Government Department of Health; updated 2024.